Eremotherium eomigrans
Quick Facts
Common Name: Wandering giant ground sloth
About two dozen skeletons of Eremotherium eomigrans were recovered from ancient lake sediments represented by the Haile 7C and 7G fossil sites.
The mounted skeleton on display at the Florida Museum of Natural History is a composite primarily consisting of bones from two similar-sized individuals.
Two very different size groups are represented by full grown adult individuals, presumably representing sexual dimorphism. In most mammals in which this is the case, the males are larger than the females, but in living tree sloths, females are slightly larger than males. So it is therefore unclear which size group represents which sex in Eremotherium.
Age Range
- Early to Middle Pleistocene Epochs; late Blancan to middle Irvingtonian land mammal ages
- About 2.6 to 0.6 million years ago
Scientific Name and Classification
Eremotherium eomigrans De Iuliis and Cartelle, 1999
Source of Species Name: The species name derived from the Greek word for ‘early’, eon, and the Latin word for ‘migrant’, migrans. The name refers to the likelihood that this species was the first of its genus (and family) to migrate to North America from South America.
Classification:Mammalia, Eutheria, Xenarthra, Pilosa, Tardigrada, Megatherioidea, Megatheriidae, Megatheriinae
Alternate Scientific Names: none
Overall Geographic Range
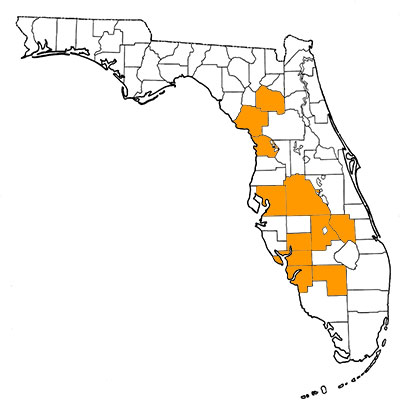
This species is known mainly from Florida. Fields et al. (2011) reported specimens from Dorchester County, South Carolina. Given its South American origins, specimens should eventually be recovered from Central America and all along the Gulf Coastal Plain. The type locality is Haile 7C, Alachua County, Florida (De Iuliis and Cartelle, 1999).
Florida Fossil Occurrences
Florida fossil sites with Eremotherium eomigrans:
Discussion
Xenarthra is the order of mammals that includes armadillos, anteaters, and sloths, including the giant ground sloths of the Miocene to Pleistocene epochs. It is believed that xenarthrans branched off from the rest of placental mammals earlier than any other clade within Mammalia, and are thus a very old lineage. Major lineages within Xenarthra are also very ancient, such that the three families of ground sloths recognized in Florida—the Mylodontidae, the Megalonychidae, and Megatheriidae—had already diverged by the Paleogene Period (Hulbert, 2001). As an old group, xenarthrans are fairly distinct anatomically and are characterized by a number of unique features including 1) an extra articulation on their lumbar (and sometimes thoracic) vertebrae called xenopophyses, 2) enamel-less, often fairly homogenous, ever-growing teeth, 3) a secondary spinous process on the scapula, and 4) an articular surface between the sacrum and one or more caudal vertebrae (Hulbert, 2001).
Xenarthrans originated in South America and were distributed almost exclusively to that continent for much of their history. During the event known as the Great American Biotic Interchange (GABI), several xenarthran taxa migrated to North America before and after the formation of the land bridge between the two continents at the Isthmus of Panama 3 million year ago. The first record of xenarthrans in North America is from the late Miocene of Florida, about 9 million years ago, which preserve fossils of giant ground sloths of the families Megalonychidae and Mylodontidae. Eremotherium eomigrans is an extinct species of giant ground sloth that belongs to a third family, the Megatheriidae, that first migrated to North America in the late Pliocene or very early Pleistocene (Hulbert, 2001). Eremotherium eomigrans is the first of its family to disperse to North America (De Iuliis and Cartelle, 1999).

Eremotherium eomigrans was one of the largest species of giant ground sloth known from North America along with its close relative, Eremotherium laurillardi. Large individuals of Eremotherium may have weighed as much as 3 tons (Hulbert, 2001). Fossils of Eremotherium eomigrans are not found as widespread as the later-occuring Eremotherium laurillardi which is known from North, Central and South America in the late Pleistocene (Cartelle and De Iuliis, 1995). The former species is nonetheless known from numerous skeletons from localities such as Haile 7C and Haile 7G in Alachua County, Florida (De Iuliis and Cartelle, 1999). This species was at first only known from Florida, but specimens from Dorchester County, South Carolina, have also been attributed to Eremotherium eomigrans based on the likely Blancan age of the locality in which they were found (Fields et al., 2011). At least in Florida the temporal range of Eremotherium eomigrans is late Blancan to Irvingtonian (Ir) 2, with the first appearance of Eremotherium laurillardi in the early Rancholabrean (Ra1). There are no known fossils belonging to the genus Eremotherium from the Ir3 interval in Florida, from about 600 to 250 thousand years ago. So there is no evidence that the two species overlapped in time. Most likely Eremotherium eomigrans became extinct during the Ir2, and Eremotherium laurillardi later dispersed from South America to replace it.
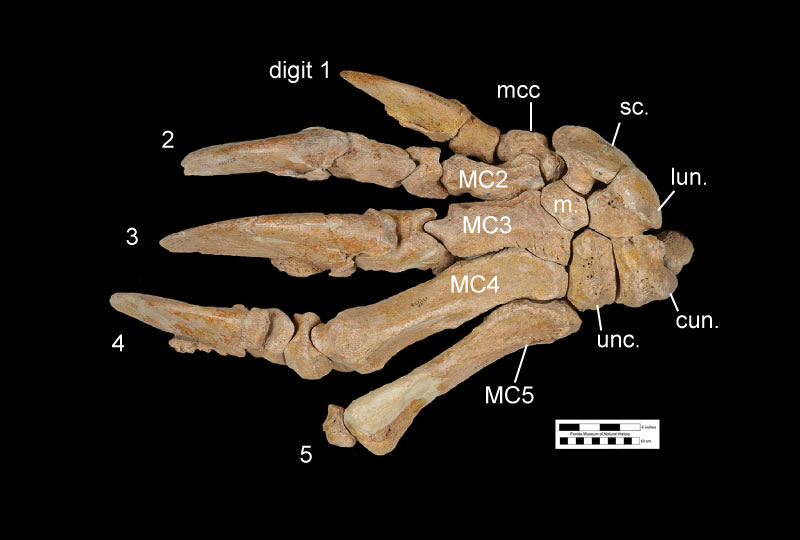
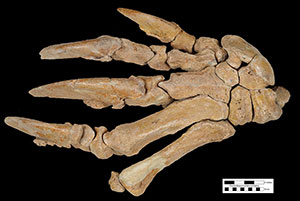
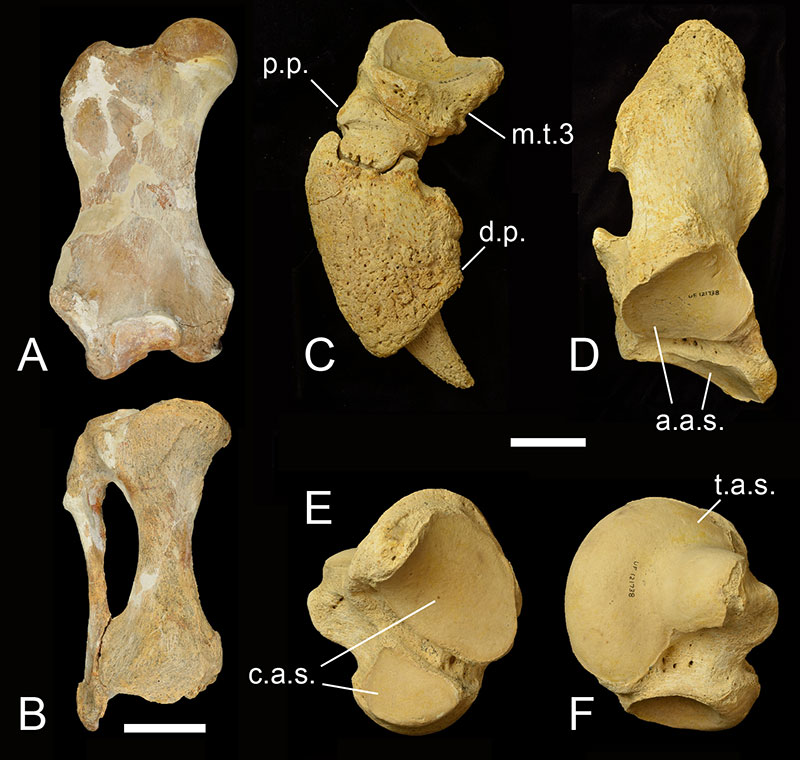
Anatomically, Eremotherium eomigrans is very similar to Eremotherium laurillardi, and the former species may be ancestral to the latter. The cranium (Figure 2) and dental anatomy of the two species is almost impossible to distinguish, and the general size and shape of many of the postcranial bones are similar between the two species and fall within the range of variation for each other (De Iuliis and Cartelle, 1999). The teeth of Eremotherium are large, rectangular when viewed occlusally, and have a unique v-shaped valley in lateral view (Fig. 2D). Both species had 5 upper and 4 lower teeth in each side of the jaw, with upper and lower crowns interlocking along the V-shaped valleys into what was likely a shearing surface for processing leaves and small branches (Hulbert, 2001). Eremotherium eomigrans in general had slightly more gracile postcranial bones (Fig. 3) and a larger lesser tubercle on the humerus than in Eremotherium laurillardi (De Iuliis and Cartelle, 1999). The major diagnostic feature of Eremotherium eomigrans is its manus (or hand; Fig. 4), which has five complete digits. Eremotherlium laurillardi has only 3 digits (they have lost digits 1 and 2) and another closely related megatheriid giant ground sloth, Megatherium americanum, has 4 digits with a loss of digit 1. While Eremotherium eomigrans is similar to Megatherium americanum in size and in some anatomical features such as more slender metacarpals, the former species is characterized by a shallower maxilla with reduced hypsodonty of the upper teeth compared to the latter species. Eremotherium eomigrans and Eremotherium laurillardi are also probably more closely related to each other than either is to Megatherium based on their shared characteristics of a rugose ectotympanic bone on the skull and the contiguous nature of the articular facet for the atlas (first cervical) vertebra on the axis (second cervical) vertebra (De Iuliis and Cartelle, 1999).
Because many specimens of Eremotherium eomigrans have been found, there is some understanding of variation within the species. Like Eremotherium laurillardi, it is thought that Eremotherium eomigrans exhibited a high degree of sexual dimorphism in body size. This is based on the size distribution of individuals, which seems to indicate that adult individuals of a larger size and of a smaller size existed within the same population (De Iluiis and Cartelle, 1999). Furthermore, Eremotherium eomigrans varied in the morphology of their wrist (carpal) and hand bones such that two variant forms can be recognized. These two forms are differentiated based on which bones are fused together in what is known as the metacarpal-carpal-complex (MCC). The MCC in one variation consists of a fusion of the metacarpal 1, trapezium, and the trapezoid bones (Fig. 5A). The second variation has a MCC composed of just the fused metacarpal 1 and trapezium (Fig. 5B). In these two variants, the shape of articulation surface of the second metacarpal with the MCC also differs based on the type of MCC the individual possessed. Evidently, individuals with these two variations in the wrist and hand bones existed at the same time in the same populations, but the first type which had an MCC with the fused trapezoid was more common in the Blancan and less common later in the Irvingtonian. However, Eremotherium laurillardi, which is thought to have evolved from Eremotherium eomigrans, had a MCC with a fused trapezoid, and lost two digits in its manus. The functional significance of these variants and the loss of fingers in later Eremotherium is unknown (De Iluiis and Cartelle, 1999).

Sources
- Original Author: Arianna Harrington
- Original Completion Date: June 5, 2015
- Editor’s Name: Richard C. Hulbert Jr. and Natali Valdes
- Last Updated On: June 8, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida