Carcharodon hastalis

Quick Facts
Common Name: none
Teeth of the extinct shark Carcharodon hastalis have been found in most Miocene and Pliocene marine deposits in Florida that produce shark teeth. It is also found in similar age deposits around the world.
Long regarded as a member of the mako shark lineage, it is now thought by many paleontologists to be ancestral to the living great white shark.
Age Range
- Early Miocene through early late Pliocene Epoch
- About 20 to 3 million years ago
Scientific Name and Classification
Carcharodon hastalis Agassiz, 1843
Source of Species Name: probably derived from the Latin word for “spear”, hasta, because of the pointed shape of the teeth, similar to a spear tip.
Classification: Chondrichthyes, Elasmobranchii, Euselachii, Galeomorphi, Lamniformes, Lamnidae
Alternate Scientific Names: Oxyrhina hastalis, Isurus hastalis, Cosmopolitodus hastalis, Isurus xiphodon, Cosmopolitodus xiphodon, Carcharodon plicatilis
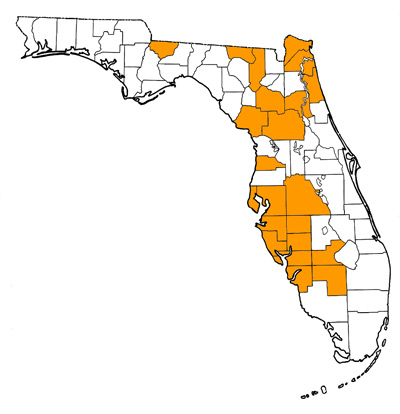
Overall Geographic Range
The species is known world-wide from marine deposits of Miocene and Pliocene age. Fossils have been found in Europe (including records in Denmark, Netherlands, Germany, Austria, Slovakia, Portugal, and Italy); Azores; United States (New Jersey, Maryland, Virginia, North Carolina, South Carolina, Georgia, Florida, and California); Mexico (Baja California Sur, Yucatan); South America (Argentina, Peru, Chile); Australia; Fiji; and Asia (Japan, India, and Taiwan). The type locality is in Switzerland (Purdy et al., 2001).
Florida Fossil Occurrences
Florida fossil sites with Carcharodon hastalis:
Discussion
Teeth of the extinct shark Carcharodon hastalis have been found in most Miocene and Pliocene marine deposits in Florida that produce shark teeth. It is typically found along with teeth of Carcharocles megalodon, Hemipristis serra, Carcharias taurus, Negaprion brevirostris, several species of the genus Carcharhinus, and several species of the genus Galeocerdo. In this assemblage, teeth of Carcharodon hastalis are the second largest, typically ranging from 1 to 3 inches (2.5-7.5 cm) in height, being exceeded in size only by those of Carcharocles megalodon.
The upper teeth of Carcharodon hastalis are broadly triangular with smooth, unserrated cutting edges (Figs. 2-4), while the lower teeth are narrower and have deeper root notches (Kent, 1994). Teeth from juveniles can have a single, very small cusplet on each shoulder. The teeth of juvenile Carcharocles megalodon and adult Carcharodon hastalis overlap in size, but the two can be easily distinguished by the lack of serrations on the cutting edges of the teeth in Carcharodon hastalis (present on teeth of Carcharocles megalodon and Hemipristis serra). Another difference is that teeth of Carcharocles megalodon have a region above the root with exposed dentine called a bourlette while teeth of Carcharodon hastalis lack such a feature, as is true of most other sharks. Fossil teeth of Isurus oxyrinchus, the shortfin mako shark, are common in Florida only on the northeast Atlantic coast (Duval and Nassau counties). Like Carcharodon hastalis teeth, they do not have serrated cutting edges, but differ in their much narrower, more curved crowns and longer root lobes (Kent, 1994).

Another large shark with smooth-edged tooth crowns that coexisted with Carcharodon hastalis was Parotodus benedeni. The latter has relatively short, thick crowns with relatively large root. Parotodus benedeni is very uncommon in Florida, with the only two known teeth coming from Duval County (Jacksonville area). The most common shark found as a fossil in Florida with a smooth-edged (non-serrated) tooth crown is the lemon shark, genus Negaprion. Negaprion teeth are smaller than those of adult Carcharodon hastalis, reaching a maximum height of 1.5 inches, with most specimens ranging from 0.5 to 1 inch. Negaprion teeth also have relatively narrower crowns, with the root being much wider than the base of the crown, and the base of the root is nearly straight or with a slight angle (Kent, 1994).


Several aspects of the classification and taxonomy of this species are currently under debate by researchers. Therefore, one will see many different names for it in scientific publications and on websites. One area of contention is into which genus to place the species. Traditionally, among members of the family Lamnidae, it was considered more closely related to the living mako sharks (Isurus paucus and Isurus oxyrinchus) than to the great white (Carcharodon carcharias) or porbeagle (Lamna nasus) sharks (Fig. 5A). This arrangement was also part of the hypothesis that Carcharodon carcharias was more closely related to megalodon and other gigantic extinct species with serrated teeth than it was to “Isurus” hastalis or any of the living species of Isurus (e.g., Applegate and Espinosa-Arrubarrena, 1996; Purdy et al., 2001; and Gottfried and Fordyce, 2001). Over the past decade, an alternate arrangement has gained favor with most researchers: that Carcharodon carcharias is not closely related to megalodon, but is instead more closely related to, and most likely the descendant of, “Isurus” hastalis (Fig. 5B; Muizon and DeVries, 1985; Nyberg et al., 2009; Ehret et al., 2012). This evolutionary transition is thought to have occurred in the eastern Pacific during the late Miocene, based on the presence there of a morphologically intermediate species, Carcharodon hubbelli Ehret et al., 2012. Workers favoring this hypothesis are divided among those who place “Isurus”hastalis in an extinct genus Cosmopolitodus (e.g., Ávila et al., 2012; Cook et al., 2010) and those who place it in Carcharodon along with the modern great white (e.g., Ehret et al., 2012). The classification used here follows Ehret et al. (2012).
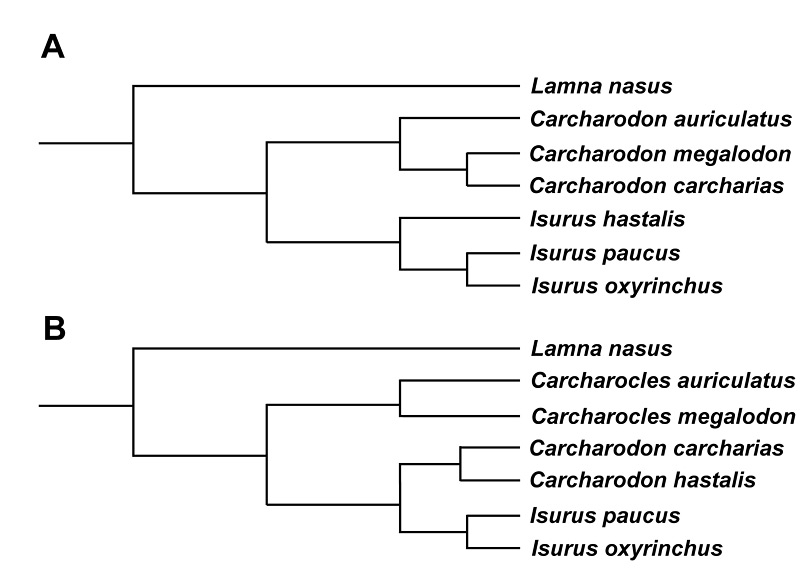
A second on-going debate is about whether or not the fossil teeth traditionally assigned to Carcharodon hastalis belong to one or two species (regardless of which genus is applied to the species). Retaining just a single species for these fossil teeth was favored by Cappetta (2006), and this interpretation is followed here. The recognition of two species was formally proposed by Purdey et al. (2001), and has been followed by some researchers, such as Aguilera and Rodrigues de Aguilera (2004), Marsili (2008), Whitenack and Gottfried (2010), and Cione et al. (2012). Under this hypothesis, even after accounting for differences in tooth width between upper and lower teeth, there is one species with relatively narrower teeth and another with relatively broader teeth. The former is more common in early to early late Miocene deposits and the latter in latest Miocene and Pliocene beds. According to Purdy et al. (2001), the narrower-toothed form retains the hastalis species name, and is thus the “true” Isurus hastalis or Cosmopolitodus hastalis, depending on one’s interpretation of lamnid evolution. The broader-tooth species was listed as Isurus xiphodon by Purdy et al. (2001) and as Carcharodon plicatilis by Cione et al. (2012), both species originally named by Agassiz in the mid-1800s.
Sources
- Original Author(s): Alexis Rojas
- Original Completion Date: December 1, 2012
- Editor(s) Name(s): Richard C. Hulbert Jr. and Natali Valdes
- Last Updated On: February 26, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida