Cormohipparion ingenuum
Quick Facts
Common Name: Noble Hipparion
Cormohipparion ingenuum was the first species of horse to be named from Florida.
Given its long, narrow muzzle and relatively short-crowned teeth, Cormohipparion ingenuum most likely fed on a mix of browse and green, fresh grasses.
During the late Miocene, Cormohipparion ingenuum was one of the more common horses in Florida. At most fossil sites of that age, it is regularly one of the three most abundant species of horse.
Age Range
- Late middle to late Miocene Epoch; early Clarendonian (Cl2) to early Hemphillian (Hh2) land mammal ages
- About 11 to 6 million years ago
Scientific Name and Classification
Cormohipparion ingenuum (Leidy, 1885)
Source of Species Name: Not explicitly stated in Leidy’s (1885:33) original description. The species name is the neuter form of the Latin word ingenuus meaning of “noble character”.
Classification: Mammalia, Eutheria, Laurasiatheria, Perissodactyla, Hippomorpha, Equoidea, Equidae, Equinae, Hipparionini
Alternate Scientific Name: Hippotherium ingenuum, Hipparion ingenuum, Nannippus ingenuus, Hipparion gratum
Overall Geographic Range
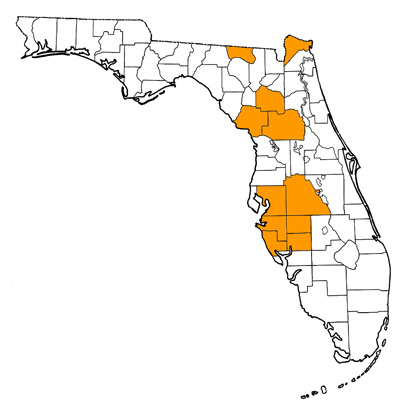
In addition to numerous records from Florida, the species is also known from the Lapara Creek Fauna of the Texas Coastal Plain (Goliad and Bee counties) and the Rancho Lobo Fauna of Honduras (Hulbert, 1988). Possibly known from the Tehuichila Fauna of Hidalgo, Mexico (see below). The type locality is Mixson’s Bone Bed, Levy County, Florida (Leidy, 1885; MacFadden, 1984).
Florida Fossil Occurrences
Florida fossil sites with Cormohipparion ingenuum:
- Alachua County—Gainesville Creeks Fauna (including Cofrin Creek, Gainesville High School Creek, Sharktooth Ravine, and Veterans Hospital Site); Haile 16C; Haile 19A; Haile 21A (reworked); Love Site; McGehee Farm; Pareners Branch Site
- De Soto County—Peace River 1
- Hamilton County—Occidental Phosphate Mine
- Hardee County—Charlie Creek 1; Fort Green Mine; Hardee Complex Mine (C.F. Industries); Hickory Creek 1; Hickey Branch Site
- Hillsborough County—Four Corners Mine; Leisey Shell Pit 1C; Kingsford Mine
- Levy County—Mixson’s Bone Bed
- Manatee County—Four Corners Mine; Port Manatee
- Marion County—Crystal Springs Mobil Homepark; Withlacoochee River 4A; Withlacoochee River 4X
- Nassau County—St. Marys River
- Polk County—Fort Meade Mine (Gardinier); Agricola Road Site, Hookers Prairie Mine; Nichols Mine Stream Matrix Locality; Grey Zone, Phosphoria Mine
- Sarasota County—lag deposit below bed 11 in Quality Aggregates Shell Pit 8A
Discussion
Cormohipparion ingenuum was the first species of horse to be named from Florida. The species has been referred to many genera over the years. It was initially placed in Hippotherium, a genus first named from Europe, but in the late 1800s it was also widely used for North American hipparion horses (e.g., Cope, 1889). In the 1900s, most paleontologists favored the use of Hipparion instead of Hippotherium, and the species was routinely placed in this genus until 1940 (e.g., Gidley, 1907). In 1940, Nannippus was raised to generic rank and began to be used for smaller North American species previously assigned to Hipparion (e.g., Stirton, 1940). So between 1940 and 1988 the species was usually called Nannippus ingenuus (e.g., MacFadden, 1984). Finally, in 1988, it was assigned to the genus Cormohipparion, and made the type species of a new subgenus, Notiocradohipparion (Hulbert, 1988). So its full, formal scientific name is Cormohipparion (Notiocradohipparion) ingenuum (Leidy, 1885).

Since it was first named by Leidy (1885), Cormohipparion ingenuum has generally been considered a valid species, regardless of which genus it was placed in. The lone exception was a few years in the late 1800s when it was regarded as a junior synonym of a hipparion species from Nebraska, Pseudhipparion gratum, first by Cope (1889) and then by Lucas in Leidy and Lucas (1896). As more specimens of this species were found, the differences between the two species were apparent and the two were no longer thought to be the same species. Indeed they are no longer considered to be in the same genus. One potential senior synonym of Cormohipparion ingenuum is “Nannippus” montezuma. It is a late Miocene species from Mexico that was named by Leidy on the basis of a single tooth two years before he named Cormohipparion ingenuum. A second, similar specimen of“Nannippus” montezuma was figured by Carranza-Casteñada and Espinosa-Arrubarrena (1994). The enamel patterns on the occlusal surface of the two teeth from Mexico can be matched with Florida specimens referred to Cormohipparion ingenuum, and the two are similar in terms of size and crown height. But before the two can be synonymized, we must learn more about the Mexican species, especially its mandibular symphysis, lower cheek teeth, and development of the facial fossa, a depression on the side of the skull in front of the eye.
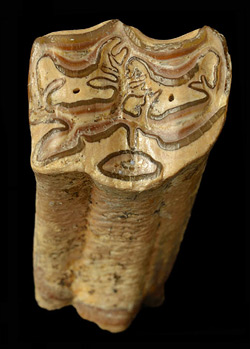
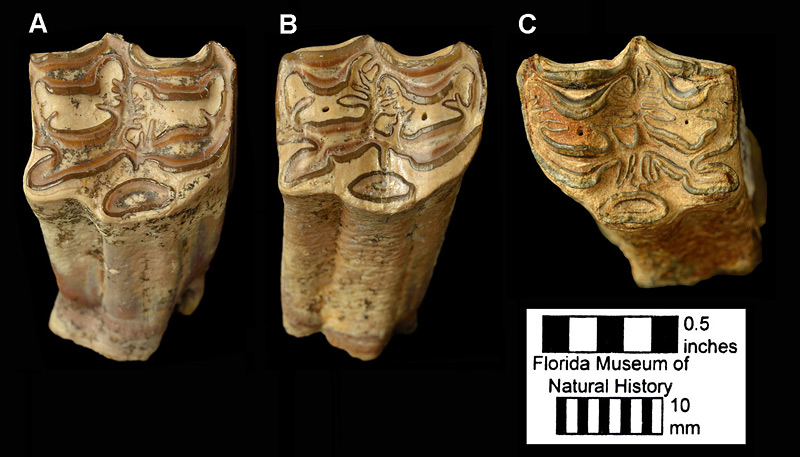
As described by Hulbert (1988), Cormohipparion ingenuum is medium-sized for a North American hipparion horse, with toothrow lengths generally in the range of 112-125 mm (ca. 4.5 inches). The complexity of enamel folds on the cheek teeth is moderate to high, but less than in Cormohipparion emsliei (Figure 3). The muzzle is very long, with the distance between the last incisor and the first cheek tooth (second premolar) equal to about 75% of the toothrow length. During the late Miocene in Florida, Cormohipparion ingenuum is often found together with a second species of Cormohipparion, Cormohipparion plicatile. These two species are very similar in tooth enamel morphology, and single teeth or small samples of teeth can be difficult to assign to one or the other species with confidence. Lengths and widths of teeth and bones of Cormohipparion ingenuum are on average about 12% smaller than those of Cormohipparion plicatile, and, in an unworn state, the molars are about 9 mm (0.35 inches) shorter in Cormohipparion ingenuum. The facial fossa on Cormohipparion ingenuum is shallower and has a less distinct edge or rim than the one on Cormohipparion plicatile. Finally, Cormohipparion ingenuum has an enamel fold on the metaconid of the lower second premolar, both deciduous and permanent, that projects anteriorly, sometimes connecting with the paraconid (Figure 4). This feature is absent on these teeth in Cormohipparion plicatile.

Given its long, narrow muzzle and relatively short-crowned teeth (for a late Miocene horse), Cormohipparion ingenuum most likely fed on a mix of browse and green, fresh grasses. Other horses of the time, most notably within the genera Neohipparion and Calippus, show adaptations characteristic of horses for which grasses comprise more than half the diet. Stable carbon isotope analysis of tooth enamel from Cormohipparion ingenuum cannot determine the relative proportion of grass in its diet, however, because grasses had the same proportion of stable carbon isotopes as shrubs and trees at the time Cormohipparion ingenuum lived in Florida. However, such analysis of stable carbon isotopes does show that all equids from the Love Site, including Cormohipparion ingenuum, lived in relatively open habitats, in contrast to the tapirs, gomphothere, rhinos, and camelids (Feranec and MacFadden, 2006). Hoffman (2006) conducted a mesowear study on the teeth of some fossil equids from Florida. In grass-eating species, teeth wear relatively flat (low relief) and the cusps rapidly wear down and become blunt. Hoffman found that in a sample of Pleistocene Equus teeth, 100% had low relief and 73% had blunt cusps, signs of a pure grass diet in a dry environment. For three species of horse from the Love Site, Cormohipparion ingenuum had the fewest teeth with low relief (50%) and blunt cusps (15%). In contrast, 75% of the teeth of Neohipparion trampasense had low relief and 41% had blunt cusps. From this it can be inferred that the percentage of grass in the diet of Cormohipparion ingenuum was less than in Neohipparion trampasense, at least within these co-existing populations.
During the late Miocene, Cormohipparion ingenuum was one of the more common horses in Florida. At most fossil sites of that age, it is regularly one of the three most abundant species of horse. The youngest records of the species are at the Withlacoochee River 4A and 4X sites, in the Hemphillian 2 interval, about 6.5 million years ago. It is succeeded by Cormohipparion emsliei, which first appears about that time, and then persists until about 2 million years ago. Because both of these species have relatively long chronologic ranges, they are not particularly useful for biochronology.
Sources
- Original Author(s): Richard C. Hulbert Jr.
- Original Completion Date: October 17, 2013
- Editor(s) Name(s): Richard C. Hulbert Jr. and Natali Valdes
- Last Updated On: December 16, 2020
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida